
O treinamento intenso ativa a síntese de proteínas, mas apenas se a nutrição certa estiver lá para apoiá-lo. Se você leu o artigo que John Meadows e eu escrevemos chamado Maximize Protein Synthesis, ou se você leu algo que a T Nation vem dizendo sobre o assunto há anos, provavelmente está familiarizado com o conceito de "janela anabólica" e a importância de nutrição peri-treino.
O que acontece no nível celular nas horas após o treinamento é preditivo de ganhos a longo prazo. Aproveite esta "janela anabólica" e você crescerá como nunca antes. Sempre perdi e, bem, boa sorte.
Obter os macronutrientes certos na hora certa é a chave, mas as macros são apenas uma parte do quadro geral. Um aspecto importante, embora muitas vezes esquecido, da síntese de proteínas musculares é o volume celular. O volume celular não é apenas cosmético; é o principal impulsionador do transporte de aminoácidos, também trabalhando nos bastidores para ativar a síntese de proteínas e suprimir a degradação de proteínas.
Um músculo cheio / volumizado é um músculo anabólico. Embora saibamos há mais de 20 anos que o inchaço celular inibe a degradação de proteínas e estimula a síntese de proteínas em certas células (1-3), até recentemente, o mecanismo que liga o volume celular à síntese de proteínas era um mistério.
O que sabemos agora é que a síntese de proteínas é controlada pela enzima mTOR, que é ativada por estresse mecânico, fatores de crescimento e leucina.
Embora todos os três sejam importantes para o estímulo de treinamento, a sinalização mTOR também depende do volume da célula.(4) Isso é especialmente importante no músculo esquelético, onde a volumização celular ativa a síntese de glicogênio e proteína e inibe a degradação de proteínas.(5, 6)
O avanço científico que levou à conexão entre o volume celular e a síntese de proteínas ocorreu em 2005, quando um grupo de cientistas descobriu que é preciso mais do que apenas leucina para ativar o mTOR - a glutamina também é necessária.(7)
Isso foi uma surpresa. Embora a glutamina seja considerada um aminoácido "condicionalmente essencial" que limita a degradação de proteínas durante traumas graves ou estresse, ela nunca foi associada à ativação de mTOR.
A glutamina era necessária para a captação de leucina e a volumização celular, ambas necessárias para ativar a síntese de proteínas. Os autores passaram a mostrar que a glutamina celular esgotamento não só resulta em uma redução do volume celular, mas também reduz a capacidade da leucina de ativar a síntese de proteínas.(7)
Essa descoberta foi enorme, porque forneceu uma ligação direta entre a glutamina, a volumização celular e a síntese de proteínas. Pela primeira vez, foi demonstrado que a glutamina é necessária para a ativação da síntese protéica pela leucina.
Embora este estudo tenha sugerido que a glutamina é uma peça muito importante do quebra-cabeça que liga o volume celular e a síntese de proteínas, o mecanismo exato não foi descoberto até 2009, quando Nicklin et al. descobriram que a exportação de glutamina está associada à importação de leucina e ativação de mTOR.(8)
Para obter leucina na célula, há um período inicial de "carga de glutamina.”Isso também puxa a água, aumentando o volume da célula. Após a fase de “carregamento de glutamina”, a glutamina é exportada para fora da célula em troca da importação de leucina.
Nicklin et al. também descobriram que os níveis de glutamina celular são limitantes da taxa para a ativação da síntese de proteínas pela leucina. Quando as células foram tratadas simultaneamente com glutamina e uma mistura de EAA contendo leucina, a ativação da síntese de proteínas foi retardada por 60 minutos. Quando essas mesmas células foram "pré-carregadas" com glutamina, a síntese de proteínas foi ativada 1-2 minutos após as células terem recebido leucina.
Este resultado foi importante porque explicou o tempo de latência para a ativação da síntese de proteínas pela leucina neste modelo experimental.
Esses resultados finalmente lançam alguma luz sobre a maquinaria celular que regula o transporte de aminoácidos e como ele está acoplado ao controle da síntese de proteínas.
Este trabalho deve ser interpretado com certo cuidado, no entanto. Uma advertência importante para esses estudos foi que eles foram realizados in vitro (i.e. em culturas de células) onde a regulação ou síntese de proteínas é muito mais simples. As células musculares são capazes de produzir glutamina conforme necessário a partir de outros aminoácidos, e a "depleção de glutamina" neste modelo de cultura de células não é representativa de mais situações fisiológicas in vivo.
Altas taxas de síntese de proteínas não podem ser mantidas indefinidamente no tecido muscular com ou sem suplementação de glutamina. A glutamina pode, no entanto, ser usada para apoiar estrategicamente a síntese de proteínas, otimizando a volumização celular durante o período pós-treino.

As células estão muito ocupadas e existem muitos canais iônicos ligados à membrana e proteínas transportadoras que regulam o tráfego para dentro e para fora da célula. Existem duas classes de transportadores de aminoácidos em particular que são importantes aqui: os transportadores de aminoácidos “Sistema L” e “Sistema A” estão mais intimamente ligados à sinalização de mTOR e síntese de proteínas.(8-10)
A atividade dos transportadores do Sistema A e do Sistema L é acoplada, o que permite que a leucina e os outros BCAAs sejam absorvidos pela célula.(11) Os transportadores do Sistema L são responsáveis pelo influxo de leucina e os outros BCAAs em troca do efluxo de glutamina.
Os transportadores do sistema A funcionam por meio de um mecanismo diferente, no entanto, onde a glutamina é acoplada à absorção de sódio.(12, 13) O acoplamento entre a absorção de sódio e os transportadores de aminoácidos do Sistema L / Sistema A é denominado Transporte Ativo Terciário (TAT para abreviar). É a TAT que, em última análise, leva a leucina para dentro da célula, levando à ativação do mTOR e à síntese de proteínas.(11)
Você pode ver como o TAT funciona na figura abaixo:
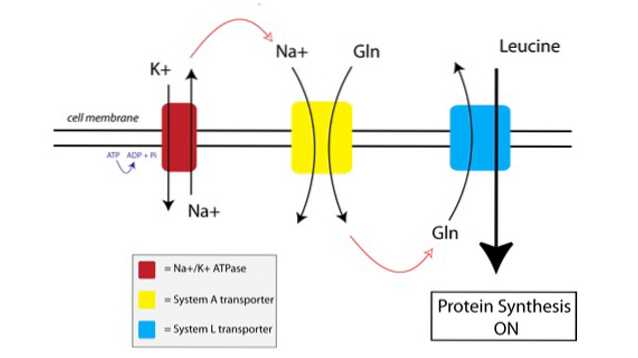
Primeiro, uma bomba ligada à membrana chamada bomba de sódio-potássio ATPase (Na + / K + ATPase, vermelho na figura acima) usa a energia do ATP para mover o sódio para fora da célula, contra seu gradiente de concentração.
O aumento da concentração de sódio fora da célula é acoplado à importação de glutamina pelo transportador do Sistema A (amarelo na figura). O influxo de glutamina e sódio na célula também puxa o excesso de água, fazendo com que a célula inche. Isso coloca a célula em um estado anabólico, preparando a maquinaria de síntese de proteínas para ativação.
Quando a glutamina atinge níveis suficientemente altos dentro da célula, os transportadores do Sistema L (em azul na figura) são ativados, o que leva a glutamina para fora da célula em troca da captação de leucina. A entrada de leucina na célula é o gatilho para a síntese de proteínas.
Embora esta tenha sido uma grande lição de bioquímica até agora, a descoberta da TAT não é importante apenas para biólogos celulares. Agora que sabemos como o volume celular está acoplado ao transporte de aminoácidos e à síntese de proteínas, podemos projetar várias estratégias nutricionais para maximizar o processo quando ele é importante - durante o período crítico de pós-treinamento.
A síntese de proteínas é totalmente dependente da hidratação celular - mesmo se você estiver um pouco desidratado, a capacidade de recuperação de um treinamento intenso está totalmente comprometida. Conseguir muita água é uma tarefa óbvia aqui, mas só água não é suficiente.
Eletrólitos como sódio, potássio, cloreto e fosfato também funcionam como "osmólitos" porque atraem água para a célula. Após uma sessão de treinamento intenso, precisamos de água, aminoácidos e eletrólitos para maximizar o processo de volumização celular que impulsiona a síntese de proteínas.
Sódio, magnésio, cálcio, potássio, fosfato e cloreto (para citar alguns) são todos importantes aqui. Em um nível mínimo (a menos que você esteja fazendo isso sob o conselho de seu médico), não se intimide com o sódio pré ou pós-treinamento. Se você está com depleção de sódio, sua bomba de treinamento será quase inexistente, e o sódio é necessário para a captação de glutamina.
Para eliminar as suposições, Surge® Workout Fuel e Plazma ™ são projetados com proporções de eletrólitos ideais para apoiar o volume celular e a síntese de proteínas.
A absorção de glutamina pela célula causa a volumização celular, preparando as células musculares para a síntese de proteínas. Como mencionado, um músculo cheio / volumizado é um músculo anabólico. Junto com a condução do transporte de aminoácidos, a volumização celular também aumenta a síntese de glicogênio e inibe a degradação de proteínas.(4-6)
A síntese de proteínas é suprimida pela depleção de glutamina, o que tem grandes implicações para atletas de treinamento pesado. Depois de uma sessão de treinamento intenso, uma resposta inflamatória é montada, o que permite que as células imunológicas trafeguem para o tecido muscular debilitado para iniciar o processo de reparo / reconstrução.(14)
A glutamina é tão rapidamente absorvida pelas células imunológicas que é considerada o “combustível do sistema imunológico.”(15) Não surpreendentemente, foi mostrado que o treinamento intenso causa depleção de glutamina no plasma.(16-18)
Por esta razão, as necessidades de glutamina aumentam no período pós-treino, onde a resposta imune local pode estar competindo pela disponibilidade de glutamina para preparar as células musculares para a captação de aminoácidos e síntese de proteínas.
Pré-carregamento células com glutamina também podem reduzir potencialmente o "tempo de latência" associado à ativação da leucina da síntese de proteínas. Se você ainda não estiver fazendo isso, tome 10-15g de glutamina ou peptídeos de glutamina imediatamente após o treino. Como os BCAAs são outro substrato favorito para a síntese de glutamina muscular e também demonstrou aumentar a produção de glutamina muscular (19-21), os BCAAs e a leucina também são úteis durante o período pré-treino para ajudar a maximizar a produção de glutamina endógena.
Nota do Editor: Todo o protocolo Plamza / MAG-10, com sua nutrição pré-treinamento, treinamento e pós-treinamento, atende às recomendações do autor para carregar os músculos com BCAAs e leucina.
Recentemente, foi descoberto que o consumo de EAAs aumenta a expressão dos transportadores de aminoácidos do Sistema A e do Sistema L.(9) É importante ressaltar que isso acontece no "nível pós-transcricional", o que significa que no nível da síntese de proteínas, onde os mRNAs existentes são traduzidos em proteínas.
Compare isso com “de novo”Expressão de proteínas - onde pode levar 16 ou mais horas para sintetizar, processar e transportar novos mRNAs - a ativação pós-transcricional da síntese de proteínas pode ocorrer em minutos a horas, permitindo que as células aumentem rapidamente o nível de proteínas específicas quando necessário.
De repente, temos mais incentivos para ter um plano de nutrição peri-treino sólido - a ingestão de EAA durante os períodos pré e peri-treino compensa muito no pós-treino, aumentando a expressão dos transportadores de aminoácidos, preparando a célula para a absorção máxima de aminoácidos e ativação da síntese de proteínas.
Proteínas isoladas ou hidrolisadas de rápida absorção, como Mag-10® Protein Pulsing Protocol ™ ou Plazma ™ durante os períodos pré e peri-treino são ideais aqui.
A insulina é o hormônio mais anabólico do corpo. Junto com a ativação direta da síntese de proteínas, a insulina também aumenta a translocação dos transportadores de aminoácidos do Sistema A para a membrana celular.(22)
Isso significa que a insulina faz com que mais transportadores do Sistema A sejam exibidos na membrana celular, prontos para conduzir mais glutamina para dentro da célula. Mais glutamina leva a mais volume celular, o que leva mais leucina para dentro da célula, levando a uma maior síntese de proteínas.
Enquanto os EAAs aumentam o expressão de transportadores AA, é o sinal de insulina que permite que eles sejam exibidos na superfície da célula, prontos para transportar novos aminoácidos para dentro da célula.
Esta é outra razão pela qual os carboidratos pré e peri-treino são uma boa ideia, a menos que você esteja no modo de perda extrema de gordura: a insulina aumenta a capacidade de transporte celular de aminoácidos.
Os carboidratos aumentam os níveis de insulina, mas certos aminoácidos também podem ser usados para potencializar a liberação de insulina. A glutamina é um poderoso ativador dos hormônios “incretinas”, que tornam as células produtoras de insulina no pâncreas mais sensíveis à glicose.(23) A glicina também potencializa a liberação de insulina por meio de um mecanismo diferente.
Embora os carboidratos pós-treino por si só aumentem os níveis de insulina, a combinação desses aminoácidos potenciadores da insulina com os carboidratos vai sobrecarregar seu pâncreas para uma liberação ainda maior de insulina. Embora seja bom manter os níveis de insulina baixos na maior parte do tempo, o aumento dos níveis de insulina no período peri-treino maximiza o transporte de aminoácidos, o volume celular e a síntese de proteínas, ao mesmo tempo que suprime a degradação de proteínas.
O tipo de treinamento anaeróbio intenso necessário para construir cargas de músculos de qualidade leva a uma produção considerável de lactato, diminuindo o pH muscular. Isso leva à fadiga muscular precoce e fraqueza, mas também certos transportadores de aminoácidos, incluindo o Sistema A, são inibidos por baixo pH.(13)
Quando o pH do músculo está baixo, a captação de aminoácidos é reduzida, o que suprime a ativação de mTOR da síntese de proteínas.(24) Também foi demonstrado que a inibição do baixo pH dos transportadores de aminoácidos do Sistema A aumenta a degradação de proteínas.(25)
É aqui que entra a beta-alanina. Os níveis aumentados de carnosina muscular atuam como um tampão de ácido natural, estendendo o limiar anaeróbio ao limitar a diminuição do pH muscular do treinamento.
A beta-alanina tem outra função importante: ajudar a manter a síntese protéica e colocá-la online mais rapidamente após um treinamento intenso, evitando a atenuação do transporte de aminoácidos.
Para aumentar os níveis de carnosina muscular, tome dois comprimidos de Beta-7 ™ três vezes ao dia.
Durante sessões de treinamento intenso, a síntese de proteínas é reduzida e a degradação de proteínas é ativada. Isso é inevitável para qualquer levantador de treinamento pesado. No entanto, até que ponto podemos minimizar os efeitos catabólicos do treinamento e quanto mais rápido podemos retornar ao "modo anabólico" durante o período pós-treinamento, em última análise, determina quão eficientemente iremos recuperar - e crescer.
O momento certo dos macronutrientes é importante, mas é um meio para um fim. O volume celular é o principal impulsionador do transporte de aminoácidos e da síntese de proteínas. Ao compreender como o transporte de aminoácidos acontece e como ele é regulado pelo volume celular, podemos colocar mais leucina nas células musculares destruídas mais rapidamente, alimentando assim o fogo anabólico e, por fim, levando a melhores ganhos.
As estratégias acima são eficazes, práticas e são baseadas nas pesquisas científicas mais recentes. Use-os como um modelo para levar seu progresso de treinamento para o próximo nível.
Aguardo suas perguntas no LiveSpill!



Ainda sem comentários